Na fase S ocorre a duplicação do material nuclear da célula. Uma célula eucariótica típica tem dois genomas, ou seja, dois conjuntos inteiros de informação genética do organismo. Um dos genomas provém do pai, e o outro, da mãe. Por isso, a célula é dita diploide, ou seja, sua ploidia é 2n.
Sse o conteúdo de DNA da célula for 2C (a unidade C é arbitrária), ao final da fase S ele será 4C, e assim as duas células-filhas receberão, cada qual, exatamente a mesma quantidade de DNA contida originalmente na célula-mãe.
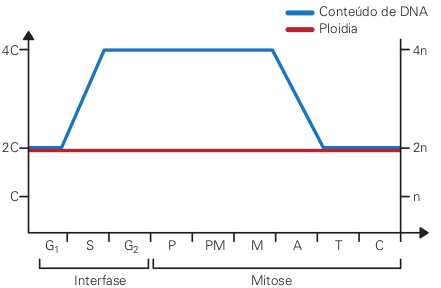
Além da mesma quantidade, elas receberão a mesma qualidade de DNA da célula-mãe, ou seja, o conteúdo de DNA, na mitose, é repartido igualmente entre as células-filhas. Elas serão, portanto, diploides, tal qual a célula-mãe.
Para a replicação do DNA, a molécula deve estar em sua fase de descondensação máxima. Em outras palavras, o DNA deve estar distendido, permitindo que enzimas afastem as duas hélices e, como vimos, dupliquem a molécula. Além de duplicação do DNA, ocorre na interfase a contínua produção de todos os constituintes celulares.
Mitocôndrias e cloroplastos se duplicam autonomamente, pois possuem DNA próprio. Também há intensa atividade de transcrição e tradução na interfase.
Nessa fase é possível que ocorram, espontaneamente, erros de duplicação do material genético. Esses erros podem ser significativos para o futuro da linhagem celular, pois a alteração de uma única base nitrogenada pode implicar a perda irremediável da informação no trecho de DNA afetado. Observe na figura abaixo um erro possível na duplicação do DNA.
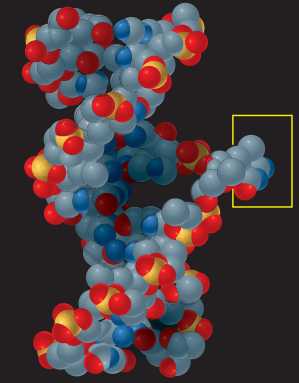
Existem diversas bases nitrogenadas, além das quatro que formam o DNA, e é possível que algumas delas sejam incorporadas a uma das fitas em formação. Como vimos, cafeína, xantina e hipoxantina são bases nitrogenadas e eventualmente podem se “intrometer” no DNA.
Essas alterações são detectadas por enzimas de reparo que monitoram constantemente as moléculas de DNA do núcleo e promovem a substituição da base nitrogenada errada pela base correta. Isso é possível, pois a base complementar do pareamento guarda a informação da base do pareamento original.
Por exemplo, em certo ponto da molécula de DNA pode haver um par citosina-hipoxantina, par não usual. A enzima de reparo tem o poder de inserir um grupo amina (NH2) na hipoxantina, que se
converte, assim, em guanina. A presença de citosina em um dos lados do pareamento garante que se restabeleça o pareamento original. Esse mecanismo de reparo é bastante eficiente.
Uma fita única, em caso de algum dano, teria a informação original irremediavelmente perdida. Além disso, a geometria da hélice dupla torna evidentes os erros de duplicação da molécula. Observe novamente a figura anterior e veja como uma base nitrogenada “intrometida” (no caso, a xantina) mudou a forma da dupla hélice, evidenciando o erro.
As lesões de pareamento de nucleotídeos que causam distorções na geometria da molécula de DNA são rapidamente reparadas por enzimas que removem a parte da cadeia anormal, gerando uma lacuna por alguns momentos. Essa lacuna é reconhecida por enzimas que atuam preenchendo-a com o pareamento normal, restabelecendo a sequência original do trecho da molécula.
Esse sistema de reparo de DNA é extremamente comum nos seres vivos, em arqueas, bactérias e eucariotos. Essas distorções da geometria molecular são facilmente induzidas por raios ultravioleta da luz solar, e, no caso humano, podem induzir a formação de tumores de pele, se ocorrerem falhas nesse sistema de reparo.
É assim interessante perceber como o DNA constitui uma molécula apropriada tanto para armazenar informação como para preservá-la quando ela é copiada, e restaurá-la, quando ocorrem problemas. A configuração em dupla fita facilita o processo de reparo, pois mantém a memória dos pareamentos.
Células com grandes danos no DNA emitem sinais capazes de paralisar o ciclo celular. Caso as enzimas de reparo não consigam deter a emissão desses sinais, a célula pode ter seu ciclo celular paralisado permanentemente, entrando no chamado G , ou sua morte pode ser induzida por apoptose.
O DNA duplicado da célula que se prepara para a divisão começa a se condensar e forma cromossomos duplicados, ou seja, com duas cromátides-irmãs. Em certo momento da mitose, os cromossomos atingem estado de condensação máxima, e ficam bem visíveis, como mostram as imagens da figura abaixo. Nessa forma, condensados, os cromossomos estão inativos e não produzem RNA.

Acima um esquema e foto de cromossomo com duas cromátides- -irmãs. Na foto (à direita), o cromossomo foi submetido a tratamento que remove algumas proteínas responsáveis por sua integridade. A ilustração (à esquerda) traz uma representação esquemática do mesmo cromossomo com a identificação das partes de uma cromátide.
Os cromossomos duplicados serão separados e divididos equitativamente pelas células-filhas, que receberão uma cromátide de cada um deles, sendo assim idênticas à célula-mãe em termos de quantidade e qualidade do DNA. As células-filhas herdam os mesmos cromossomos da célula da qual se originaram.
Veja também
- Divisão celular
- Métodos de estudo da célula
- As Bases Nitrogenadas – nucleotídeos
- Ácido Nucleico: o que são, os tipos, resumo
- Estruturas Celulares e Organelas citoplasmáticas